The commercial types have been systematically selected for degenerative micro sports
Where is this documented?
The commercial types have been systematically selected for degenerative micro sports
Where is this documented?
Larry,
Many years ago, while I was at K-State, I went through Modern Roses V marking all the varieties that had originated as sports. I then wrote the names of varieties and sport-parents on slips of paper so that I could sort them by “family”. I didn’t have a personal computer back then (they didn’t exist yet) so the work was rather tedious but satisfying. Long story short, the sporting tendency does seem to run in families. ‘Peace’, for example, is descended from Pernetianas as well as from ‘Ophelia’. ‘Talisman’ (Ophelia x Souv. de Claudius Pernet) also gets a double dose of sportiness.
Selection for vigor was well demonstrated by Breese, Hayward and Thomas (1965) Somatic selection in perennial ryegrass. Heredity 20: 367-379.
“An experiment has shown that the rate of tillering (= asexual reproduction) in perennial ryegrass can be significantly altered by selection within clones. The response to this somatic selection depended on the age of the clone such that response was obtained in young clones raised from seedlings but not in clones with a long history of uninterrupted asexual propagation. The response was also dependent on the genotype of the clone and there was some evidence to suggest that the somatic lability of genotypes was predisposed by a history of adaptation to asexual propagation previous to the sexual cycle.”
http://bulbnrose.x10.mx/Heredity/King/VegetativeSelection.html
Then there’s this demonstration of rebloom vs. vigor in a rose:
Brownell: What is an “Everblooming” Cllmber? (1944)
“While the rose hybridizer cannot successfully combine the true reblooming quality with the ordinary cane-growth of climbers, very satisfactory types may be produced by encouraging the vigorous branching growth of the flowering stems. An illustration of this is the variety Orange Everglow in which these two types of blooming habit are present and segregated. Certain confirmation lies in the fact that if the once-blooming cane-growth is not removed it may by its vigor smother and prevent the establishment of reblooming wood.”
http://bulbnrose.x10.mx/Roses/breeding/BrownellClimber1944.html
Karl
Yeah, I’m not challenging the assertion I am just hoping that there might be some documentation other than pedigrees.
I can’t provide documentation, but I can mention a theory known as “Muller’s Ratchet” that apparently had a broad acceptance before epigenetics, transposons, RNAi and related phenomena became widely known.
Proceedings of International Workshop on Sweetpotato …, 2001
D.R. La Bonte, A.Q. Villordon, and D.S. Fajardo.
Mutations in Sweetpotato
“Muller’s ratchet (Muller, 1964) was a convenient working model for what we initially hypothesized in sweetpotato (Villordon and La Bonte, 1995, 1996). Muller states, “species without a sexual phase are impeded in evolution but also subject to genetic deterioration”. Muller’s “ratchet mechanism” embodies the concept that mutations can only increase in time. In other words, a chance mutation rarely reverts; additional mutations accumulate and result in lowered “fitness”, or from our perspective, clonal decline.”
The authors went on to discuss transposons, changes in heterochromatin, methylation and other things.
“Sweetpotato is unique in that new propagules from fleshy roots arise adventitiously from callus, wound periderm, vascular cambium, or anomalous cambium (Edmond and Ammerman, 1971; Esau, 1977; Fahn, 1982). We hypothesize that this process is akin to regenerating plants from callus culture — a prime mechanism for promoting genetic instability.”
Some root-sports have been reported in roses: ‘Happenstance’ and ‘Little Mermaid’ from ‘Mermaid’, ‘Baby Jane Clare’ from a seedling of ‘Sequoia Ruby’ and ‘New Dawn’ from ‘Dr. W. Van Fleet’.
‘Chevy Chase’ is reported to sport to a pink flowered form when severely pruned, presumably due to the formation of adventitious shoots. ‘CC’ may be a chimera.
Many years ago I rooted cuttings of some Polyanthas of the “Koster” group that I was growing, with a plan to take root cuttings in hopes of finding more sports. I got side-tracked and never took up the effort again.
Karl
I am totally confused. Years ago when I first started posting on this forum, I mentioned the use of Pernetiana cultivars (Foetida Hybrids). The response I received from various members was “don’t use them there full of BS and you will introduce BS into your seedlings”. Well that outlook has certainly changed looking at this thread.
Well I did use Pernetiana’s in my breeding, but only those which showed minimal BS infections. The use of these Pernetianas was not to obtain yellows but to enrich certain colours making them more brighter.
You don’t really have to use Foetida to obtain clear bright yellows, these seedlings which germinated this year obtained this yellow even in our bright Ozzie sunlight with high UV levels.



From your post Warren, I have taken this bit,
You don’t really have to use Foetida to obtain clear bright yellows, these seedlings which germinated this year obtained this yellow even in our bright Ozzie sunlight with high UV levels.
Could you explain please if you could, which plants you used to achieve your colours.
Regards David.
I will speak freely about my species hybrids, but these I won’t go into to much detail as I know of a couple of major European breeders which browse this forum. I hope every one gets what I mean.
Warren, I quite understand what you are driving at in your comment, also in your crosses ( I think). One interesting thing I should point out about your 1st and 3rd pics, is the red styles. This is characteristic of many, but not all yellows coming all the way from the very early Pernetianas. The point being you probably have the same set of genes that would be found in a cross to Persian Yellow, but with a different general genetic background. The earlier discussion was, if I recall correctly, driven by Kim’s comments (paraphrase) that we don’t need to reinvent that wheel. But more recently it is about the possibility of bringing in garden shrub architecture from the likes of PY or AC or AY along with the yellow intensity, while using selected seed parents to avoid bringing in the BS susceptibility that is at least purported to come from those 3.
Personally, I’m more interested in understanding the genetics going on here so I’m chasing down both paths, starting direct for AC for its brilliant color, and making use of the most disease resistant yellows I can get my hands on. Right now that is Carefree Sunshine, Sunny KO and Rainbow KO. But none of those are really a good rich yellow and they fade badly. To me it looks like the richest yellows frequently come with red styles or filaments or both. I think this is telling us that some of the rich yellows actually contain a trace of anthocyanin, even in the petals. Too much and they look orange. Take a look at Shock Wave on HMF. Strong yellow, fades to puny pink after a week or so, especially in cool climate. Pinking is not obvious in warmer conditions, just the fading.
I think what we are dealing with is not simple old-fashioned Mendelian genetics of one gene = one character. Instead we have a basic pathway to yellow, and a bunch of regulatory regions and factors that ramp up or down the expression of genes for the enzymes that make the yellow color. Some of those switch on in pollen and filaments giving bright yellow. Others switch off giving white pollen and green filaments. In a like manner anthocyanins are turned on and off, though genes for the enzymes are present in all the flower parts. So you can get petals with no, or almost no, red, and a ton of yellow, while having anthers, filaments, stigmas and styles that are red + yellow in various combinations. AC has red stigmas and inner petal surface, while AY has neither. PY is pretty solid yellow in many photos, but not all; some show red styles.
There isn’t a Foetida-based yellow which doesn’t fade within the first hour or so in Zones 8b through 10a here in Southern California. Yellow, in particular, is short lived here to begin with, but none of the Pernetianas I have grown or observed in these conditions lasted a full afternoon in any season. Perhaps in a more northerly climate they may hold their color, but definitely not here. They held precisely to Dr. Nicholas’ statement that they were “thirty minute roses”, as they were faded and usually blown in about that length of time. Don’t misunderstand, I love them and collected every one I could lay my hands on all those years ago, but their blooms were extremely “fleeting”.
Thanks Larry
Thanks Kim
I was going through the genetics of the parent in relation to the three pics above, they have around 10% R. foetida either through direct crossing with R.foetida or by its hybrids , through the Pernetiana lines.
Harkness said if your going to breed yellow look at the stamen colour, but when you look at it, a lot of petals are converted stamens.
warren
Harkness said if your going to breed yellow look at the stamen colour
Hmmm. Can you elaborate, Warren?
Possibly as Ralph Moore also said, “stained yellow” versus “pure yellow”. All potential flower colors appear in all the plant parts. If you want pure yellow, choose a yellow with yellow stamen and without a red blush to the petal reverses, a red “stain” or sunburn. Like, to stabilize mauve, a stained yellow will produce a stained mauve, like Paradise or Angel Farce. An unstained yellow would produce a pure mauve without the red blush.
Don,
I don’t have Harkness’ statement, but Le Grice (1973) wrote:
“It is obvious that a working hybridist must find some method by which he can get some idea as to the purity of colour in the strain on which he is working. Two shades of deep yellow are available. The first is a mustard yellow such as Lydia [H. Robinson, 1949], which owes its intensity to its greenish tinge, showing that the red-blue factor is present and active. The other, such as Allgold, is a clear deep yellow with no green shade. By studying the whole flower one can see the difference. In the first case somewhere in the stigma, style, anther, or filament the red or green or both red and green will show. In the second case the pure colour extends through the whole flower; stigma, style, anther, and filament are of the same shade as the petal.”
http://bulbnrose.x10.mx/Roses/breeding/Legrice/Legrice1.html
Karl
Do you have any non-Foetida yellows that fade less?
Most of the yellow color in roses is due to the various carotenes that are present. In addition, some forms of quercetin and luteolin may add to the appearance.
Journal of Photochemistry and Photobiology A. 136(1-2): 87-91 (31 August 2000)
The photostabilities of naturally occurring 5-hydroxyflavones, flavonols, their glycosides and their aluminium complexes
Gerald J. Smith, Scott J. Thomsen, Kenneth R. Markham, Claude Andary and Dominique Cardon
"In aqueous methanol solution, luteolin and flavonol 3-glycosides exhibited no degradation over periods of up to 15 h of UV irradiation. However, the flavonols studied were all found to degrade and their relative photostabilities correlate with their redox potentials. Quercetin was the least stable. In the presence of aluminium ions, all the flavonoids, including luteolin, were degraded by UV irradiation.
“In contrast to the absorption spectra in dilute solution, the reflectance spectra of both quercetin and luteolin deposited on a cellulosic substrate exhibited strong absorptions beyond 400 nm. On this substrate these flavonoids displayed the characteristic yellow colour associated with flavonoids in some environments. Although the quercetin yellow faded rapidly on exposure to UV radiation, the colour of luteolin darkened. This was due to the formation of a photoproduct absorbing maximally at 450 nm.”
http://irl.cri.nz/photostabilities-naturally-occurring-5-hydroxyflavones-flavonols-their-glycosides-their-aluminium-co
Of course, what happens under laboratory conditions (methanol solution, cellulosic substrate) may not match perfectly what happens in vivo. But it is interesting to note that the presence of aluminum ions hastened the degradation by UV. This implies that the fading would be greater in acidic soil (which liberates aluminum ions) than in alkaline soil.
Also, the darkening of luteolin “due to the formation of a photoproduct absorbing maximally at 450 nm” is worth noting. I have observed that ‘Devoniensis’ becomes more yellow as the flowers age. I don’t know that this involves luteolin.
http://bulbnrose.x10.mx/Roses/Rose_Pictures/D/Devoniensis.html
http://bulbnrose.x10.mx/Roses/Rose_Pictures/D/devoniensisCl.html
Karl
As I wrote, yellow, in general, is quite fleeting. The Foetida types tend to be more intense and their fade is faster and more dramatic. Their petal substance, particularly in the earlier types, is also very soft. The flowers “blow” very quickly, even in the more double types.
Eugster and Marki-Fisher say that the flavonoids contibute little to yellow color. They also say the most UV-stable, late stage, epoxide forms of the carotenoids are the most color dense and spectrally pure ones that show up in R. foetida and it’s very close hybrids. The farther you get from the foetidas (including Persiana) the less dense the pigments and the less stable they are.
Below is a chart showing the biosynthetic pathways for carotenoid synthesis (red annotations refer the the pigments as numbered in Eugster & Marki-Fisher). I’ve posted this before, from which you should get the idea that the genetics of yellow pigmentation are highly complex with lots and lots of room for short-circuiting.
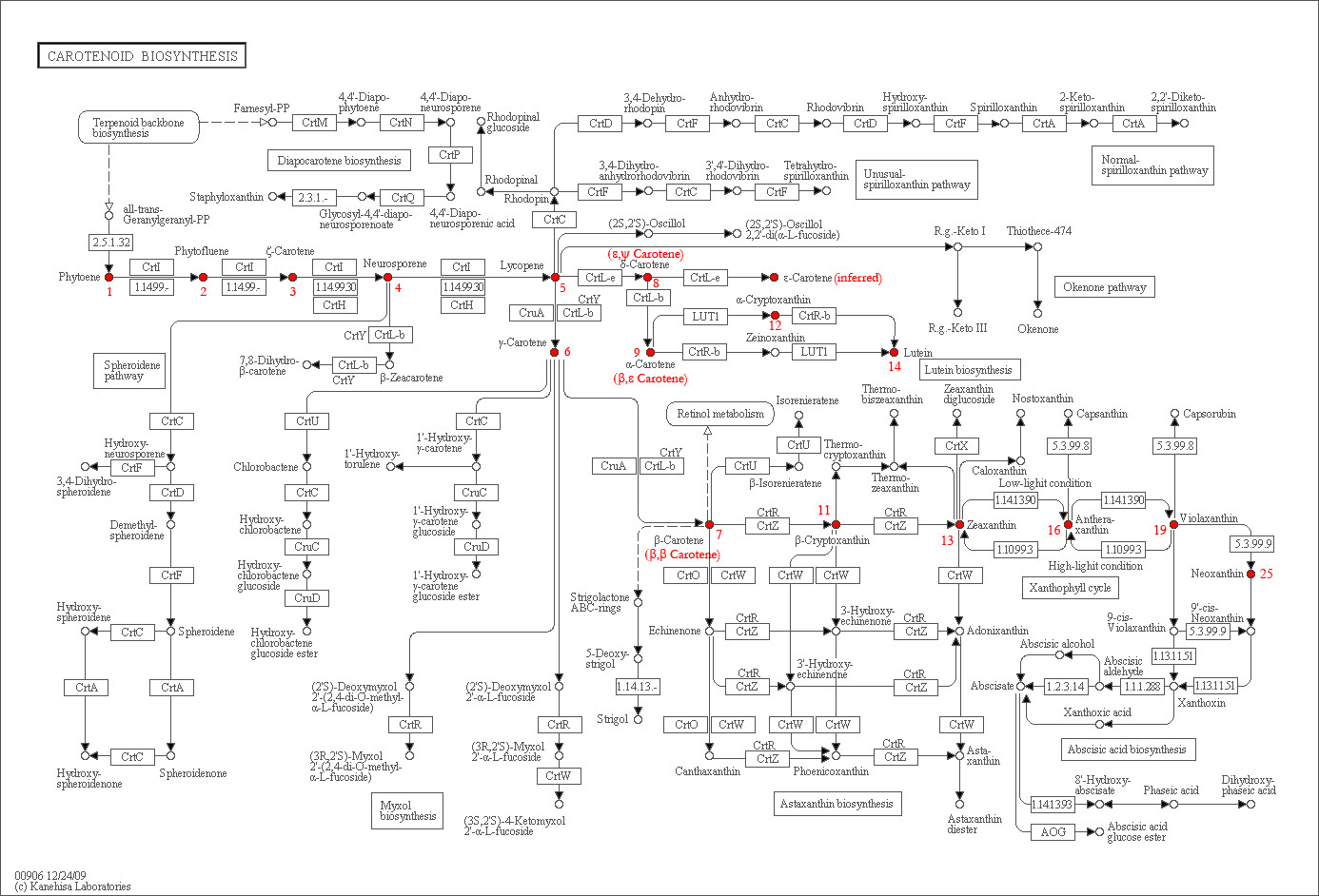
It is pretty much biologically impossible (without resort to clever genetic engineering) to have your yellow and keep it too. That is because, even when nothing interferes with their production, a pathway called the violaxanthin cycle sacrifices the late stage epoxide carotenoids to protect plants from ultraviolet light. See
“…all rose flowers, including yellow ones, contain quite a lot flavonoids. However, only when they are present in concentrations, together with carotenoids, do they contribute to visible absorption”
The work of Eugster and Märki-Fischer, quoted above, is obviously important. But they did skimp a bit on their discussions of flavonoids and anthocyanin pigments. For example, Harborne (1976) noted, “The common flavonols probably contribute to yellow flower colour when (a) they are methylated or (b) they are present in certain unusual glycosidic forms. … Quercitin 7- and 4’-glucosides have absorption spectra similar to quercetin itself and may therefore provide some yellow in gorse Ulex europeaus, in Rosa foetida and in other petals in which they occur.”
I must emphasize SOME yellow.
As for anthocyanins, Mikanagi, et al. (2000) give more information than did Eugster and Märki-Fischer.
Biochemical Systematics and Ecology 28: 887-902 (2000)
Anthocyanins in flowers of genus Rosa, sections Cinnamomeae (=Rosa), Chinenses, Gallicanae and some modern garden roses
Mikanagi, Saito, Yokoi, Tatsuzawa
Abstract: Forty-four taxa of three sections (Cinnamomeae (=Rosa) 26, Chinenses 8 and Gallicanae 10) and eight modern garden roses in the genus Rosa were surveyed for their floral anthocyanins. Eleven anthocyanins: 3-glucosides and 3,5-diglucosides of cyanidin (Cy), pelargonidin (Pg) and peonidin (Pn), 3-rutinosides and 3-ρ-coumaroylglucoside-5-glucosides of Cy and Pn, and Cy 3-sophoroside, were isolated from flowers of these taxa and identified by chemical and spectroscopic techniques. Four anthocyanins: Cy 3-rutinoside, Pn 3-rutinoside, Pn 3-ρ-coumaroylglucoside-5-glucoside and Cy 3-sophoroside were found for the first time in Rosa flowers.
Investigated sections of wild roses showed characteristic distribution of anthocyanins. Cy 3,5-diglucoside was the dominant anthocyanin detected in all three sections, but accumulation of Pn 3,5-diglucoside distinguished sections Cinnamomeae from other sections, and the occurrence of Cy 3-glucoside separates section Chinenses from others.
Cy 3-sophoroside was detected in large amount in some taxa of section Cinnamomeae: e.g., R. moyesii and its related cultivars, and R. rugosa cv. Salmon Pink. The acylated Cy glycoside was found in all sections and also in some modern garden roses, while the acylated Pn glycoside was detected in the section Cinnamomeae, but not in sections Chinenses and Gallicanae. According to anthocyanin distribution patterns, eight groups were classified chemotaxonomically in genus Rosa.
…In particular, the blood-red coloured flowers of R. moyesii cv. Geranium contained [cyanidin 3-sophoroside] as 56% of total anthocyanins. This pigment is probably associated with the special colour of that cultivar.
And another:
Harborne and Wiliams: Anthocyanins and other flavonoids. Nat. Prod. Rep., 2001, 18: 310-333
The discovery of cyanidin 3-sophoroside is of biosynthetic interest, since the related flavonol 3-sophorosides are dominant copigments in rose petals.
Eugster and Märki-Fischer mentioned that they found peonin in R. moyesii, but not a word about the more interesting cyanidin 3-sophoroside.
Karl
Great paper, thank’s for the citation. I found it online and will chew on it for a while.
I’m intrigued by the moyesii-specific glycosides. I have a number of F1 moyesii’s and am working to produce F2’s. I built a reflectance spectrometer a while back with the thought of using it for screening hybrids. It’s been on the shelf for lack of some good reference data to work with. Maybe this paper will give me what I need for distinguishing cyanidin sphorosides.
Incidentally, my foundation moyesii breeder looks to my eye like it actually has peonidin rather than cyanindin. I did have Geranium at one time (but lost it to a dreadful anther smut of some sort) and the color of that was the same as my pure species. There is a Superba out there somewhere which I tried once to bud unsuccessfully. I should probably get it this spring, I think Andy van Cleve carries it.
Could one use Chelsea filter like theory to isolate different pigments by eye.
Chelea’s are the wrong wavelengths from what I can tell just looking online.
Someone posted here a while back about about using gel lens filters, dunno what became of that but we had a discussion about which filters were better so you could find it with the search function. It depends, of course, on the pigments you want to detect.
Ah, found it.
http://www.rosebreeders.org/forum/viewtopic.php?f=2&t=54339&p=55560&hilit=filter#p55180